Annelida
29/10/2013 22:29Grupo: Marcela Akemi, Mayara Oliveira, Roberta Bonela e Thabata Tailer
Annelida
O filo Annelida ocupa o grande nicho ecológico dos vermes em ambientes marinhos, de água doce e terrestre. Há aproximadamente 16500 espécies descritas. O nome do filo deriva do latim annulatos e compreende os vermes segmentados, cada segmento apresentando partes internas e externas repetidas, o que é conhecido como homologia seriada, ou seja, são estruturas que apresentam a mesma origem e que evoluíram da mesma forma resultando na metameria.
São vermes eucelomados e esquizocélicos, ou seja, a mesoderme origina-se na parte do arquêntero próximo ao blastóporo, onde uma massa de células prolifera-se na blastocele e em seguida se divide deixando uma cavidade no seu interior, o celoma. Cada segmento apresenta um par bilateral de cavidades celômicas.
São animais bilaterais, ou seja, o lado esquerdo e direito são espelhos, a metade dorsal é diferente da ventral, assim como a metade anterior é diferente da posterior.
O corpo dos anelídeos é composto por três regiões: o prostômio, o tronco e o pigídio. Na região anterior localiza-se o prostômio, uma região pré-segmentada, onde se encontra o cérebro e as estruturas sensoriais. O pigídio encontra-se na extremidade posterior do corpo e é onde está o ânus. Tanto o prostômio quanto o pigídio não são considerados segmentos por não se originarem na zona de crescimento da larva trocófora. Esta zona de crescimento no indivíduo adulto restringe-se a uma região imediatamente a frente do pigídio. O crescimento do corpo é caracterizado como teloblástico e é resultante da adição de novos segmentos na parte posterior, sendo o segmento mais novo o anterior ao pigídio e o segmento posterior ao prostômio o mais antigo.
Os metâmeros são separados uns dos outros por septos intersegmentares. Alguns anelídeos apresentam segmentação homônima, ou seja, os segmentos são muito parecidos entre si. Há também algumas espécies em que o padrão de segmentação é heterônomo, que é caracterizado por grupos de segmentos que apresentam especializações em diferentes funções.
A parede do corpo dos anelídeos apresenta uma cutícula colágena fibrosa. Algumas espécies marinhas tubícolas (poliquetas) não apresentam cutícula, mas os tubos que secretam assemelham-se a esta. Há também a presença de cerdas que são estruturas constituídas por quitina que se projetam da epiderme e promovem a tração e outras tarefas.

O sistema nervoso central desse filo é composto por um cérebro anterior dorsal e um par ventral de cordões nervosos. Em cada segmento os cordões nervosos possuem um par de gânglios unidos entre si por um nervo transversal (comissura).
Os anelídeos utilizam estruturas sensoriais que encontram-se distribuídas na cabeça, apêndices e outras regiões do corpo. Algumas espécies podem apresentar também uma grande variedade de órgãos dos sentidos como ocelos, olhos, estatocistos e órgãos nucais quimiorreceptores. Geralmente os órgãos sensoriais ocorrem no prostômio, peristômio e em alguns segmentos anteriores.

Lumbrineris sphaerocephala cabeça ventral (esquerda), parapódio, e cabeça dorsal
O fluído celômico funciona como esqueleto hidráulico contra o qual os músculos agem para alternar a forma do corpo. Os músculos que compões os segmentos são os longitudinais que ao se contraírem fazem com que o fluído celômico exerça uma força direcionada lateralmente e que o corpo se amplie, já os músculos circulares quando contraídos fazem com que o fluído celômico exerça uma força direcionada anterior e posteriormente resultando no alongamento do corpo. A tração com o substrato é aumentada pelas cerdas pareadas e quitinosas que estão posicionadas lateralmente em cada segmento.
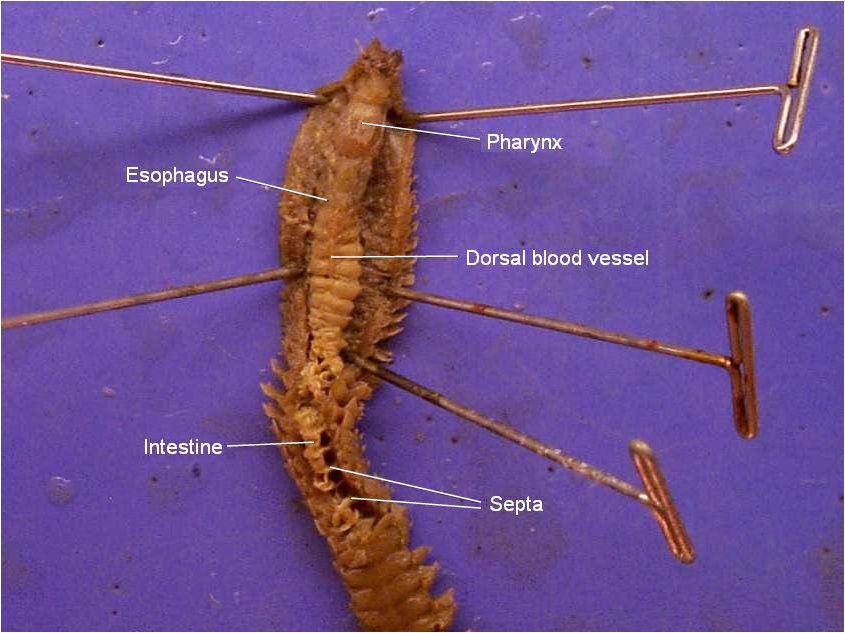
Os anelídeos possuem um tubo digestivo que vai da boca até o ânus. O intestino fica suspenso dentro do celoma por meio de mesentérios longitudinais e dos septos, através dos quais o intestino penetra. A digestão é extracelular.
O sistema sanguíneo é desenvolvido e pode ocorrer em pequenos vasos e em vasos maiores. Os vasos principais são o vaso sanguíneo dorsal e o vaso sanguíneo ventral. O sangue flui em direção ao anterior no vaso dorsal e posterior no ventral.
Os órgãos excretores deste filo são protonefrídeos. O túbulo nefridial de cada nefrídeo penetra no septo e passa para o segmento posterior antes de se abrir para o exterior do corpo.
O sistema nervoso é formado por uma massa ganglionar dorsal anterior, um par de conjuntivos anteriores circundando o intestino e um cordão nervoso ventral único ou duplo com um inchaço ganglionar e nervos laterais em cada segmento.
São primitivamente animais gonocóricos que liberam seus gametas através dos seus metanefrídeos . Uma vez liberados, a fecundação ocorre externamente na água do mar dando origem posteriormente a larvas trocóforas. Apresentam todas as formas de reprodução assexuada: fragmentação, brotamento, paratomia e estolonização.
As espécies de anelídeos estão distribuídas em duas divisões: Polychaeta e Clitellata, sendo esta última dividida em Oligochaeta e Hirundinoidea.
1)Polychaeta
A classe polychaeta é composta de cerca de 8 mil espécies. A maioria escavadora, mas alguns rastejam sobre o substrato ou sobre organismos a ele aderidos, outros perfuram conchas e rochas, muitos podem nadar quando necessário e alguns são exclusivamente pelágicos.
Essa classe é constituída, geralmente, por vermes de tamanho mediano com cerca de 10 centímetros de comprimento. Alguns vermes intersticiais medem menos de um milímetro, enquanto outros podem medir mais de um metro de comprimento (Eunice, Nereis e Polydontes). O maior Polychaeta conhecido é Eunice aphroditois que mede mais de 3 metros de comprimento.
- Estrutura
Esses anelídeos possuem pernas que recebem o nome de parapódios. Os parapódios são projeções laterais da parede do corpo que podem ser complexas e grandes, ou pequenas e pouco desenvolvidas que são encontrados em pares em cada segmento. Um parapódio é basicamente birreme, ou seja, composto de uma parte superior, o notopódio, e outra ventral, o neuropódio. Os cirros são processos sensoriais em forma de tentáculos que são projetados da base dorsal do notopódio e da base ventral do neuropódio. Os lobos parapodiais (ramos) possuem bolsas chamadas sacos setígeros que secretam os feixes de cerdas. As cerdas podem assumir formas variadas e os feixes setígeros de uma determinada espécie podem apresentar mais de um tipo de cerda. Essas estruturas são utilizadas na locomoção, aumentando a tração com o sedimento ou superfícies.
Os élitros são estruturas modificadas dos cirros dorsais em escamas e estão presentes em várias famílias de poliquetas errantes, coletivamente chamadas de vermes-escamosos. Como a maioria vive em lugares apertados, a escama proporciona um canal protetor contra a corrente ventilatória. Os élitros também portam várias estruturas sensoriais, como no verme-escamoso (Aphrodita), que toda sua superfície dorsal é revestida por um feltro piliforme, composto de cerdas, que surge dos notopódios e corre para trás na superfície dorsal do animal.

A epiderme do poliqueta é composta por uma única camada de epitélio cuboide ou colunar, que é coberta por uma cutícula colágena, fibrosa e fina. Um componente comum do epitélio são as glândulas secretoras de muco. Por baixo do epitélio, existe, em ordem, uma camada de fibras musculares circulares, uma camada mais espessa de fibras musculares longitudinais e uma camada fina de peritônio.
Os poliquetas são os únicos entre os anelídeos que apresentam diversos tipos de apêndices cefálicos sensoriais. Nos apêndices prostomiais estão incluídos antenas anteriores e palpos ântero-ventrais. Normalmente os cirros tentaculares sensoriais ou palpos tentaculares são encontrados no peristômio de muitos poliquetas que possuem função alimentar.
O pigídio possui um par de cirros pigidiais, mas em alguns poliquetas, são encontrados mais de um par. No entanto, poucos poliquetas apresentam essa estrutura, porque diferentes estilos de vida dessa classe levaram a graus variáveis de modificação no plano básico.
As formas cavadoras e tubícolas podem apresentar uma forma diferenciada dos segmentos. O tronco se especializou em regiões distintas (tórax e abdome como resultado da variação na forma dos parapódios, ou presença ou ausência de brânquias. O número de segmentos pode variar de menos de 10 a mais de 200.

Poliqueta sabelídeo branchiomma em costão rochoso
1.2)Sustentação e Locomoção
Os Poliquetas podem se locomover de várias formas diferentes, tudo depende das adaptações do corpo e estilo de vida. Uma forma de locomoção é a escavação peristáltica, muito comum nas famílias de poliquetas que possuem corpos alongados, parapódios e apêndices da cabeça reduzidos. Normalmente a musculatura circular é desenvolvida e os septos completos, fazendo com que o fluido celômico se restrinja a cada segmento. Exemplos de famílias que se locomovem assim são: Arabellidae, Lumbrineridae, Captellidae e Orbinidae.
Outros escavadores possuem outros meios de locomoção, por exemplo, o verme-sanguíneo (Glycera) e o arenicola usam uma faringe eversível para perfurar e ancorar no sedimento e depois com a retração, puxar a si mesmo para frente. Alguns membros da família Nephtyidae (verme-dançarino), Armandia e Ophellidae produzem rápidas ondulações laterais do corpo e nadam através da areia fracamente consolidada. Os habitantes de sedimentos muito pequenos usam cílios para locomoção, exemplos são os políquetos das famílias Diniphilidae e dos Diurodrilidae intersticiais. Poliquetas das famílias como nereidas (Nereididae), filodicideos (Phyllodocidae) e os hesionideos (Hesionidae) rastejam sobre superfícies utilizando os parapódios e cerdas bem desenvolvidas. O movimento é resultado da ação combinada dos parapódios, da musculatura da parede corporal e do fluido celômico. Os músculos longitudinais são melhores desenvolvidos que os circulares e os septos tendem a ser incompletos.
1.3)Hábitos
Os poliquetas podem ser errantes (movendo-se livremente) ou sedentários, mas nem sempre a distinção é precisa. Os poliquetas errantes incluem diversas espécies pelágicas, algumas que rastejam por baixo de conchas e rochas, algumas que são escavadoras ativas na areia e na lama, mas também ocupam tubos estacionários. Muitas espécies sedentárias assim como o Phragmatopoma caudata, constroem e vivem em buracos, galerias ou tubos. As espécies tubícolas obrigatórias geralmente não podem deixar seus tubos e só podem projetar suas cabeças através da abertura dos mesmos.
Já os escavadores podem ser ativos, que cavam através de peristalse, ou podem ser sedentários, aonde a escavação é iniciada quando o organismo é jovem, porém o mecanismo ainda é desconhecido.
Os de forma tubícola ocupam buracos escavados em forma de “u” ou verticais simples, mais ou menos fixos. Exemplos de poliquetas tubícolas são os Arenicolidae e os Terebellidae.

Terebellidae
Ainda existem os simbióticos, no qual possui um relacionamento com outro animal, podendo ser o hospedeiro ou o hóspede. O papel do hospedeiro é exercido primariamente pelos tubícolas não-carnívoros e pelos poliquetas escavadores, que proporcionam um abrigo ventilado e protegido. Os poliquetas comensais distribuem-se em numerosas famílias por toda classe, mas os vermes escamosos (Polynoidae) incluem o maior número. O parasitismo não é comum entre os poliquetas. O Lobrorostratus e outros Oenonidae vivem no celoma de outros poliquetas e podem ser quase tão grandes quanto seu hospedeiro. Os ectoparasitas poliquetas incluem o Ichthyotomidae sugadores de sangue, que se prendem às nadadeiras das enguias marinhas. Os Mizóstomos são um grupo peculiar de poliquetas comensais e parasitários e são encontrados sobre equinodermos, especialmente crinoides.
1.4)Alimentação
Os poliquetas são classificado em raptoriais, comedores de depósitos e comedores de suspensão, que inclui os filtradores. Entretanto, há vários métodos e preferências entre cada uma destas divisões básicas.
1.4.1)Raptoriais
Os raptoriais ou predadores, incluem membros de muitas famílias de espécies habitantes da superfície, muitos grupos pelágicos, tubícolas e habitantes de galeria ativo. Suas presas são pequenos invertebrados (incluindo outros poliquetas), que são capturados por uma faringe eversível, a probóscide. O animal everte sua faringe através de rápidas contrações da musculatura da parede do corpo nos segmentos anteriores, aumentando a pressão hidrostática nos espaços celômicos e causando a eversão. A faringe possui duas ou mais mandíbulas que capturam o alimento e quando a pressão reduz-se no fluido celômico, a faringe é recolhida pelos músculos retratores, que se estendem da parede corporal à faringe. Os predadores podem ingerir material vegetal e detritos. Alguns comem restos alimentando-se de praticamente todo material morto que encontram.
1.4.2)Os poliquetas comedores de depósito direto
Alguns poliquetas comedores de depósitos diretos consomem areia ou lama diretamente aplicando a boca contra o substrato. A ingestão é facilitada por meio de uma faringe simples e não muscular. Entre os poliquetas, encontram-se nessa divisão os tubícolas e os escavadores. Os Vermes-bambu (Maldanidae) são exemplos de tubícolas consumidores de depósito. O verme vive de cabeça para baixo e ingere o substrato no fundo do tubo de grãos de areia. Os cílios dentro da faringe evertida empurram as partículas soltas no interior do intestino, e ocorre na verdade uma triagem de partículas.
1.4.3)Os poliquetas comedores de depósito seletivo
Os consumidores de depósito seletivo não tem uma faringe especializada. As estruturas especiais da cabeça se estendem sobre ou no interior do substrato. O material do depósito adere a secreções mucosas na superfície dessas estruturas alimentares e é depois transportado até a boca ao longo de tratos ou sulcos ciliados. Esses poliquetas selecionam o material de depósito orgânico dentre as partículas de areia.
Os órgãos ingestivos prostomiais dos terebelideos (por exemplo, a Amphitrite e Terebella) são formadas de grandes cachos de tentáculos contráteis, que se esticam sobre a superfície do substrato por meio de um rastejamento ciliar. Os detritos da superfície aderem ao muco secretado pelo epitélio tentacular. As partículas movem-se para baixo de uma calha ciliada formada pelo tentáculo enrolado, e o alimento acumula-se na base dos tentáculos, cada um dos quais enrolado por cima de uma pálpebra que limita a boca. Os cílios no lábio então empurram o alimento para o interior da boca.
1.4.4)Comedores de suspensão
Muitos poliquetas tubícolas e escavadores são filtradores. Alimentam-se de partículas que se aderem à superfície das estruturas alimentares (processos que coletam detritos e plâncton da água circundante) e depois são transportadas até a boca ao longo de tratos ciliados. Nos filtradores, a estrutura é utilizada para capturar o alimento são os radíolos.
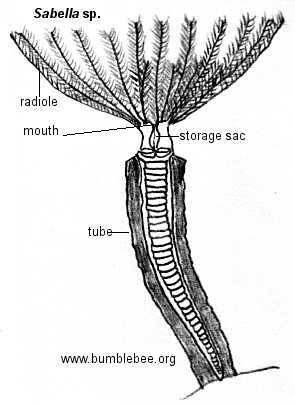
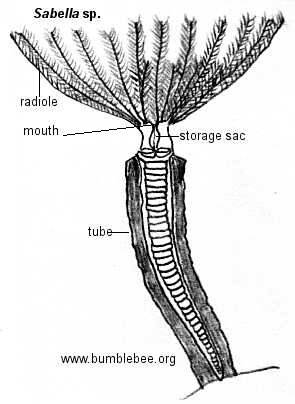
Os radíolos bipinulados em forma de coroa dos vermes-leque serpulideos, sabelideos e espirorbideos formam um funil de uma ou duas espirais quando expandido para fora da extremidade do tubo. O batimento dos cílios situados nas pequenas nadadeiras produz uma corrente de água que flui através dos radíolos no interior do funil e depois para cima e para fora. As partículas prendem-se nas pequenas nadadeiras e são empurradas pelos cílios em um sulco que corre na extensão de cada radíolo. As partículas são transportadas ao longo do sulco para baixo até a base do radíolo, onde ocorre um processo de triagem mais completo. As partículas maiores são rejeitadas e as menores são transportadas por tratos ciliados no interior da boca.
O mecanismo alimentar dos quetopterideos difere dos outros filtradores. Os Chaetopterus (que vivem em tubos com formato de “U”) tem uma estrutura corporal altamente modificada. Os notopódios no décimo segundo segmento são longos e em forma de asa e o epitélio é ciliado e rico em glândulas secretoras de muco. Dos segmentos 14 a 16, os notopódios são fundidos formando leques semicirculares que se projetam como anéis contra a parede cilíndrica do tubo. O batimento desses leques faz com que a água entre e saia do tubo por extremidades opostas. Os notopóodios pareados em forma de asa esticam-se para fora ao redor das paredes do tubo e secretam um muco entre elas. O filme mucoso é secretado continuamente por cada notopódio até assumir um formato de saco. A água que é trazida pelos batimentos dos parapódios em leque passa por essa rede de muco que puxam os detritos e o plâncton suspensos. Os objetos grandes são detectados pelos cílios peristomiais; os notopódios em forma de asa são então puxados para poder deixados objetos grandes passarem por eles. O saco mucoso carregado de alimento fica sendo enrolado continuamente em uma esfera pela cúpula dorsal. Quando a esfera atinge um determinado tamanho, a esfera solta-se dos notopódios e enrola-se coma esfera. A cúpula então se projeta para frente e doposita a esfera alimentar mucosa sobre um sulco meio-dorsal ciliado, que se estende até aparte anterior do verme, e a esfera é transportada até a boca.
1.5)Sistema digestivo
O canal alimentar dos poliquetas é um tubo reto que se estende da boca, parte anterior, até o ânus, parte posterior. O canal é dividido em faringe, esôfago curto, estômago, um intestino e um reto. Muitas vezes, a diferenciação dessas regiões só pode ser feitas quando analisadas histologicamente.
O estômago ou o intestino elaboram enzimas digestivas para a digestão extracelular. O intestino é o local de absorção, então não raramente, as paredes se dobram para aumentar a área da superfície intestinal.
1.6)Circulação e Trocas Gasosas
As brânquias são muito comuns em poliquetas, mas elas podem variar tanto em estrutura quando em localização. Elas nunca se fecham dentro de câmaras protetoras; algumas espécies, que vivem em tubos, já possuem as brânquias protegidas. As brânquias não aparecem em poliquetas muito pequenos ou que possuem longos corpos cordiniformes (tais como Lumbrineridae, Oenonidae e Capitellidae escavadores).
Nos vermes escamosos, a troca gasosa é feita pela superfície corporal dorsal que é tampada pelos élitros. Os cílios na superfície dorsal criam uma corrente de água que flui posteriormente por baixo dos élitros.
As brânquias, normalmente estão associadas aos parapódios e em muitos casos constituem partes modificadas deste. O notopódio pode possuir um lobo branquial achatado, que pode agir como uma brânquia, como nas nereidas. Muitas vezes o cirro dorsal pode encontrar-se modificado para servir como uma brânquia, ou as brânquias surgem da base do cirro dorsal.
Muitas espécies sedentárias têm brânquias aderidas às extremidades anteriores próximas a abertura dos tubos, por exemplo, as brânquias de alguns terebelideos (como a Amphitrite). Nos vermes-leque ,Sabellidae e Serpulidae, os radíolos bipinilados que compões os leques servem de locais de trocas gasosas.
A ventilação pode ser promovida por cílios branquiais ou por contrações branquiais, mas muitos poliquetas escavadores e tubícolas empurram a água através de seus tubos ou buracos por meio de contrações corporais peristálticas ou ondulatórias. De acordo com uma pesquisa conduzida por C.P. Mangum, a atividade ventilatória aumenta 15 vezes com a exigência de oxigênio do verme, mas ocorre um aumento de aproximadamente 20 vezes no consumo de oxigênio.
1.7)Transporte Interno
A circulação na maioria dos poliquetas resulta na movimentação do fluido no sistema sanguíneo vascular e no celoma. Nos vermes que possuem variação no septo, ou seja, septo reduzido, como nos vermes-sanguíneos glicerídeos, o sistema celômico substitui o sistema sanguíneo e transporta substâncias por todo o corpo. As espécies muito pequenas também não tem um sistema sanguíneo, e algumas vezes também as cavidades celômicas.
Na maioria dos poliquetas, no qual o sistema circulatório é desenvolvido, o sangue circuladentro de vasos fechados. Em um sistema sanguíneo típico, o sangue flui anteriormente em um vaso dorsal situado sobre o trato digestivo; na parte anterior do corpo, o vaso dorsal conecta-se com o vaso ventral por meio de um a vários vasos ou por meio de uma rede de vasos que passa ao redor do intestino.

Em cada segmento o vaso ventral da origem a um par de vasos parapodiais ventrais, que suprem os parapódios, a parede corporal e os nefrídeos, e a vasos intestinais ventrais, que suprem o intestino. Esses vasos se interconectam por meio de uma rede de vasos menores.
O sangue dos poliquetas está confinado em vasos de tamanhos pequenos e grandes, e em algumas espécies ocorrem seios de grande volume tipicamente na parede intestinal. As brânquias são geralmente providas com alças vasculares aferentes e eferentes, permitindo um fluxo de duas mãos. Os radíolos dos vermes-leque, além de capturarem alimento, funcionam também na troca gasosa e contém um único vaso (dentro do qual o sangue flui em marés, para dentro e para fora). Em glicerídeos, as brânquias são irrigadas com fluido celômico e não com sangue.
Em geral, o sangue é empurrado por ondas peristálticas de contração que ocorrem sobre os vasos dorsais. Muitos poliquetas têm bombas acessórias semelhantes ao coração distribuídas ao longo do sistema sanguíneo-vascular.
O fluido celômico possui mais células que o sangue. Nos poliquetas pequenos, o sangue é geralmente incolor, mas nas espécies maiores e nas que escavam em fundos macios, o sangue contém pigmentos respiratórios dissolvidos no plasma. A hemoglobina é o mais comum dos pigmentos, mas nos vermes-leque, o pigmento é a clorocruorina, que possui uma coloração verde. As moléculas extracelulares localizadas no plasma sanguíneo não só transporta oxigênio como ajuda na osmorregulação e ajudam a manter o volume plasmático do sangue. O fluido celômico circula por meio de contrações musculares da parede do corpo ou por contrações musculares e cílios no revestimento celômico.
Em alguns poliquetas, a hemoglobina pode estocar oxigênio durante períodos de repouso entre ventilação ou na maré baixa, quando a pressão do oxigênio na água do buraco e na areia reduz. Os poliquetas são oxiconformadores, ou seja, o consumo de oxigênio é regulado em parte pela quantidade de oxigênio disponível no ambiente circundante. De acordo com a pesquisa de E. Ruby e D. Fox, o escavador Euzonus mucronatus usa seu deposito de oxigênio hemoglobina para sustenta-lo durante as 4 horas de anoxia durante as marés baixas. Se for sujeito a períodos mais longos de anoxia, o verme muda para o modo de respiração anaeróbio, e pode sobreviver por até 20 dias.
1.8)Excreção
Os órgãos excretores dos poliquetas são nefrídeos filtradores, em que distribuem em pares por segmento, mas algumas famílias podem possuir poucos pares ou até mesmo um par para o verme inteiro. A extremidade anterior do tubo nefrídico situa-se no celoma do segmento imediatamente anterior àquele de onde o nefridióporo se abre. O tubo penetra no septo posterior do segmento, estende-se no interior do próximo segmento, onde pode se enrolar e depois abrir-se para o exterior na região do neuropódio.
Dependendo do formato do corpo, os poliquetas têm protonefrídeos ou sistema de metanefridicos. As famílias de poliquetas que não possuem sistema sanguíneo-vascular ou têm o sistema sanguíneo-vascular reduzido, possuem protonefrídeos. Todas as famílias restantes possuem sistema sanguíneo-vascular e metanefrídicos.
Acredita-se que a ultrafiltração (primeiro passo para produzir urina) ocorra através da parede do vaso nos poliquetas vascularizados. Depois o ultrafiltrado modifica-se e passa para o exterior através de um duto ciliado, o tubo metanefrídico. Na ausência dos vasos sanguíneos, acredita-se que a ultrafiltração ocorra à medida que o fluido celômico se desloca através das paredes das células terminais protonefrídicas. A reabsorção ocorre no túbulo protonefrídico. As células terminais protonefrídicas (solenócitos) têm flagelo único e um longo colar de filtração tubular.
O nefróstomo é um funil ciliado aberto presente na extremidade pré-septal dos metanefrídios, por exemplo, na Nereis. O nefróstomo tem um envoltório externo de peritônio, e o interior é densamente ciliado. O nefridióporo abre-se na base do neuropódio no lado ventral. O revestimento inteiro dos túbulos é ciliado.
Nos vermes-leque, no qual apresentam apenas um par de nefrídeos funcionais, os dois nefrídeos se reúnem na linha média para formar um canal mediano único, que se estende para frente para abrir-se através de um único nefridióporo na cabeça. A urina é liberada diretamente no lado externo.
O tecido cloragógeno, os celomócitos e a parede intestinal podem exercer papéis acessórios na excreção. O tecido cloragógeno é composto de células peritoneais marrons ou esverdeadas na parede intestinal ou em vários vasos sanguíneos.
1.9)Curiosidades
“Os acidentes com poliquetas são raros e ocorrem principalmente com pescadores de mexilhões, uma vez que estes são encontrados entre as colônias desses moluscos. Os poliquetas podem ferir mergulhadores com mordidas com suas mandíbulas quitinosas ou introduzir setas ou espículas. O envenenamento é basicamente irritativo, podendo ser observado parestesia parcial do membro afetado e necrose local (HADDAD JÚNIOR, 2003). “
“O inseticida Cartap , usado na agricultura, é um derivado sintético da Nereistoxina (NTX) que ocorre naturalmente na saliva de poliqueta Lumbriconereis Heteropoda. Altamente tóxica a insetos, bloqueia os receptores nicotínicos de acetilcolina da junção neuromuscular. Em vertebrados a ação é muito baixa ou inexistente, mostrando a alta especificidade do produto (DELPECH et al., 2003).”

Verme de fogo

Larva de poliqueta
Clitellata
Durante o curso da evolução, no ancestral dos Clitellatas o cérebro foi deslocado do prostômio para o tronco, diferenciando-os dos Polychaeta.
Nesta divisão estão incluídas minhocas e as sanguessugas. Os Clitellatas não apresentam parapódios, nem apêndices cefálicos e pigidiais. O que caracteriza esse grupo é a presença de um clitelo. O clitelo é uma série de segmentos que são cobertos por uma espessa epiderme glandular que forma uma cintura característica, ele engloba os gonóporos femininos e se localiza posteriormente a eles; secreta muco para cópula, albumina para nutrição dos ovos e um casulo no qual os ovos e albumina são depositados. Os clitelados são hermafroditas com cópula e suas gônadas estão situadas apenas em alguns poucos segmentos genitais, sendo os testículos anteriores aos ovários.
O desenvolvimento do zigoto ocorre no casulo e possui desenvolvimento direto.

Oligochaeta:
2.1)Estrutura
Algumas bibliografias sugerem um número aproximado de 6000 espécies e sua maioria vive em água doce ou ambientes terrestres, mas algumas espécies também se expandiram para ambientes de água salobras e marinhas. A sua grande maioria é cavadora, embora algumas habitem nas camadas de detritos epibentônicos ou vivam entre filamentos de águas. Há também uma pequena parcela que é tubícula ou parasita. Seu comprimento varia de menos de 1 mm a até mais de 3 metros. Apesar de não possuírem parapódios, geralmente apresentam poucas cerdas e alguns apêndices na cabeça.
O corpo deste grupo pode ser homônimo ou apresentar especialização como brânquias e diferentes tamanhos, formas e espessuras de cerdas. A maioria destas são móveis e utilizadas frequentemente para cavar. Em volta delas, aderidas à base, há músculos protratores e retratores que permitem às cerdas serem expostas ou recolhidas.

2.2)Parede do corpo e Celoma
Principalmente nas espécies terrestres há uma fina cutícula que recobre uma camada epidérmica contendo células glandulares mucossecretoras. Cílios ocorrem algumas vezes em certas partes da epiderme. Abaixo desta, encontra-se a musculatura circular e longitudinal, sendo a última revestida pelo peritônio. As minhocas tem os septos bem mais desenvolvidos e podem ter esfíncteres ao redor da abertura dos septos para controlar o fluxo do fluído celômico. Os septos intersegmentares geralmente são bem desenvolvidos e musculares. Cada compartimento celômico se comunica com o exterior através de poros que regulam a saída de fluído celomático para superfície corporal. Provavelmente essa saída de fluido celomático fácil ita a troca gasosa e evita a dessecação, pois mantém a película úmida.
2.3) Sustentação e Locomoção
Os oligoquetas dependem do seu esqueleto hidrostático para sustentação e locomoção. Na ausência dos parapódios (que funcionariam como remos), os oligoquetas realizam contrações peristálticas e manipulação das cerdas para se mover. Diferente dos poliquetas que perderam grande parte dos seus septos intersegmentares, a maioria dos oligoquetas os manteve completos, assim, uma constrição em uma área do corpo, não afeta outro segmento, ou seja, o fluído celômico não extravaza para outro segmento. Assim, a diminuição no diâmetro de um segmento é acompanhada pelo aumento de seu comprimento e vice-versa.
Uma onda peristáltica é gerada por uma sequência de impulsos do cordão nervoso ventral e dos neurônios motores associados. As contrações musculares circular e longitudinal modificam o segmento de longo e fino para curto e grosso gerando a onda. Entretanto, essa alternância na forma do segmentos de nada adiantaria se não houvesse as cerdas pois o corpo não produziria nenhum movimento para frente. As cerdas evitam o deslizamento para trás ao se projetarem.
Há alguns oligoquetas que utilizam a boca como um tipo de ventosa contra o substrato, atuando como as cerdas na locomoção. Outros, podem ter neurônios estimulados causando rápida contração dos músculos longitudinais em respostas de fuga rápida ou recolhimento para dentro da galeria.
2.4) Alimentação
Por não apresentarem apêndices na cabeça, isso impede a alimentação através dos tentáculos. A ausência de parapódios também elimina a possibilidade do animal ficar parado e gerar correntes de água de onde o alimento poderia ser extraído. A maioria das espécies são detritívoras, sobre tudo de matéria orgânica vegetal, pedradora ou comedora direta de depósitos.
Os oligoquetas dulcícolas se incluem nos predadores e capturam suas presas por sucção de sua faringe muscular; podem everter a probóscide onde se situam glândulas secretoras de muco, onde as presas são capturadas e recolhidas para com retração da probóscide.
Os detritívoros geralmente vivem em camadas superficiais de detritos orgânicos no fundo de lagos onde, pequenas partículas de alimentos são digeridas por ação musculas ou ciliar do trato digestivo. Também são ingeridos microorganismos vivos junto com os detritos.
A maioria dos oligoquetas terrestres e muitos aquáticos são comedores de depósitos .
As minhocas cavam através do solo, ingerindo o substrato à medida que se deslocam. Quando a terra passa no trato digestivo o material orgânico é digerido e absorvido enquanto o material inorgânico sai pelo ânus. Alguns cavadores terrestres utilizam a boca como uma ventosa na superfície do solo para obter pedaços de alimentos relativamente grandes, um exemplo são pedaços de folhas em decomposição.
2.5)Sistema digestivo
O sistema digestivo é um tubo reto, a boca localiza-se abaixo do prostômio, que leva a uma faringe bem ampla e logo após há um estreito e tubular esôfago. A parte posterior do esôfago tem regiões dilatadas formando um papo onde o alimento é armazenado e uma ou mais moelas musculares que são revestidas por cutícula na trituração do material ingerido. No esôfago, podem existir evaginações revestidas por tecido glandular, que removem o cálcio do material ingerido e são chamadas de glândulas calcíferas. Quando em excesso, o cálcio (calcita) é liberado de volta à luz do trato digestivo, onde sai do corpo pelo ânus, pois não é absorvida. Estas glândulas também tem função de regular os níveis de íons cálcio e carbonato no sangue e fluidos celômicos, tamponando o pH destes.
A digestão é principalmente extracelular. Em espécies terrestres, a área superficial do intestino é aumentada por uma região chamada tiflossole. O alimento é transportado por ação peristáltica da musculatura da parede do tubo digestivo e também por movimentos associados a locomoção.
2.6)Circulação e trocas gasosas
As trocas gasosas de quase todos os oligoquetas ocorre por difusão. Secreções das glândulas mucosas e o fluido liberado pelos poros umidecem a superfície da epiderme e facilitam essas trocas gasosas. Alguns oligoquetas apresentam brânquias especializadas .
Os oligoquetas maiores apresentam hemoglobina dissolvida no plasma sanguíneo. Em condições normais da galeria, a pressão parcial de oxigênio é a mesma daquela na atmosfera acima do solo, quando essa pressão parcial cai, a hemoglobina aumenta sua capacidade de transportar oxigênio. Após chuvas fortes, o nível de oxigênio no solo cai, fazendo com que algumas minhocas procurem a superfície para respirar. Há oligoquetas que suportam níveis relativamente baixos de oxigênio e até completa falta dele em curtos períodos de tempo.
2.7)Excreção e osmorregulação
O nefrídeo típico de oligoqueta conta com nefrostômio pré-septal, um canal curto que penetra pelo septo e um nefridioduto pós-septal, que pode estar enrolado de formas diferentes e algumas vezes, dilatado formando uma bexiga. O metanefrídeo se encontra pareado e disposto segmentarmente, em geral em todos os segmentos, exceto pelos das extremidades anterior e posterior e geralmente se encontra na região ventral do animal. Os oligoquetas aquáticos são amoniotélicos e os de forma terrestre são ureotélicos.
Os resíduos são transportados aos nefrídeos por meio do sistema circulatório e por difusão através do fluído celômico. Uma grande quantidade de reabsorção seletiva ocorre para dentro do fluxo sanguíneo eferente, facilitando a excreção eficiente, bem como equilíbrio iônico e osmorregulação.
As células cloragógenas são estruturas arredondadas e amareladas que revestem o exterior da parede fina do intestino dos oligoquetas e possui papel similar ao do fígado de animais mais evoluídos: remove os resíduos circulantes no sangue, concentra-os e os converte em amônia, ureia e creatinina, então essas excretas são dirigidas novamente para o sangue ou para o interior do celoma, aonde serão eliminados pelos nefrídeos.
Os nefrídeos são órgãos importantes para o balanço de água e sais minerais. Nos animais de água doce, o excesso de água é excretado e sais são retidos por reabsorção seletiva e ativa ao longo do nefridioduto. Já nas formas terrestres, as minhocas não são osmorreguladoras; ao invés disso perdem e ganham água de acordo com a quantidade de água disponível no meio. Sob condições normais, a conservação de água pelas minhocas é provavelmente realizada de muitas maneiras, como por exemplo a produção de ureia permite a excreção de uma urina relativamente hipertônica se comparada a de animais estritamente amoniotélicos, também pode haver tomada ativa de água e sais do alimentos, através de paredes do trato digestivo e adaptações para permanecer em ambientes úmidos.
2.8)Sistema nervoso
O sistema nervoso desses animais inclui um gânglio cerebral supra-entérico unido a um cordão nervoso ventral ganglionar por conectivos circum-entéricos e um gânglio subentérico. O gânglio cerebral em oligoquetas ocupa uma posição mais posterior do que em poliquetas devido a redução do tamanho da cabeça mas assim como em poliquetas, o par de cordões nervosos ventrais está quase sempre fundido num trato único em oligoquetas e pode conter algumas fibras gigantes e o gânglio subentérico é o centro de controle motor dos movimentos do corpo; o gânglio cerebral medeia essas atividades por influências inibitórias.
As reações iniciadas pelo gânglio subentérico causa uma série de reações de estímulo e resposta envolvendo os gânglios segmentares é responsável pleo movimento independente, mas coordenado, de cada segmento durante a locomoção. Se o gânglio subentérico for removido, os movimentos normais continuam, mas não há respostas a estímulos externos.
2.9)Órgãos dos sentidos
Exceto pelos oligoquetas dulcícolas, que apresentam ocelos pareados na extremidade anterior, basicamente todo o restante dos vermes dessa classe têm fotorreceptores simples distribuídos pela epiderme. São, geralmente, sensíveis à luz intensa e reagem de forma a se afastar da mesma.
Os órgãos sensoriais da epiderme são unidades receptoras que podem possuir terminações nervosas livres ou se agruparem em tubérculos com processos sensoriais que se projetam acima da cutícula. Estes tubérculos geralmente tem função tátil e há suspeitas que sejam quimiorreceptores – fornecendo informações sobre o meio, como mudanças no pH, localização de alimento, secreções de outros vermes. Os tubérculos formam três anéis em volta de cada segmento e são mais numerosos nos segmentos mais anteriores, principalmente no prostômio.
2.10)Regeneração e reprodução assexuada
A regeneração em oligoquetas varia muito entre as espécies. Algumas regeneram qualquer parte do corpo, enquanto outras têm uma capacidade de regeneração bem baixa.
A classe em questão apresenta uma polaridade da parte anterior para a posterior que torna a regeneração mais fácil nos segmentos posteriores do que no anteriores. Diferente dos poliquetas, apresentam um número fixo de segmentos e, durante a regeneração, nunca há a formação de mais segmentos do que havia anteriormente. Uma teoria (Moment, 1953) explica esse mecanismo de controle do número de fragmentos a partir do gradiente elétrico do corpo do verme.
Segundo Moment, cada segmento apresenta um potencial elétrico ligeiramente distinto e, quando há a perda de alguma parte do corpo, a restituição do mesmo ocorre de modo a alcançar o potencial prévio do(s) segmento(s) em questão, mantendo assim o número adequado de segmentos.
A maioria dos oligoquetas dulcícolas é capaz de se reproduzir assexuadamente. Entretanto, esse tipo de reprodução é sazonal e se alterna com a reprodução sexuada. Geralmente, ocorre no verão pois as temperaturas são mais amenas e a disponibilidade de alimento é bem maior. A maturação e reprodução sexuada ocorrem no início do outono para que os ovos eclodam na primavera.
Reproduzem-se por fissão transversal, por fragmentação (seguida pela regeneração de cada seguimento e até por brotos, onde “precursores de filhotes surgem no corpo do verme parental e, quando maduros, são liberados por fissão.
2.11)Reprodução sexuada
Os oligoquetas são hermafroditas e possuem sistemas reprodutores complexos e com gônadas permanentes. O sucesso desses animais foi resultado em grande parte da reprodução com cópula, troca de espermatozóides e desenvolvimento direto. Apesar do investimento energético ser muito alto neste tipo de reprodução, a grande quantidade de zigotos que sobrevivem compensa.
O sistema feminino é formado por um par único de ovários e se encontram posteriormente ao sistema masculino. Os óvulos são liberados no espaço celômico adjacente podendo ser armazenados em ovissacos até atingirem a maturação. Próximo ao ovissaco está um funil ciliado que leva os óvulos maduros até o oviduto que o transportará ao gonóporo.
Já o sistema reprodutor masculino apresenta um ou dois pares de testículos localizados em um ou dois segmentos específicos. Os espermatozóides são liberados no espaço celômico, onde amadurecem e são coletados pelas vesículas seminais. Quando maduros, os espermatozóides são liberados dessas vesículas e coletados pelos funis espermáticos ciliados, passando pelos ductos espermáticos até chegar aos gonóporos.
A maioria dos oligoquetas possuem receptáculos seminais, as espermatecas, que se abrem para o exterior por meio de poros.
O clitelo é uma região formada por tecido glandular em que há três tipos de células que secretam substâncias importantes na reprodução: muco para a cópula, substâncias que compõem a cápsula do casulo e albumina. A posição do clitelo e o número de segmentos envolvidos são diferentes em casa espécie. Nas formas dulcícolas, o clitelo se encontra perto dos gonóporos, já nas formas terrestres, é posterior ao gonóporo.
Durante a cópula, há o alinhamento dos corpos de dois oligoquetas em direções opostas. O muco produzido pelo clitelo mantêm os dois indivíduos nessa posição. Vários oligoquetas posicionam-se de forma a alinhar os gonóporos masculinos com as espermatecas do outro. Pode haver a presença de cerdas especiais que ajudam a manter os animais unidos. Algumas minhocas não alinham os gonóporos com as espermatecas do outro e, ao invés disso, desenvolvem goteiras espermáticas externas onde os espermatozóides viajam até a espermateca. Essas goteiras são temporárias, formadas por contração muscular e cobertas por muco. Após a troca de espermatozóides os animais se separam.
Dias após a cópula, o clitelo produz o casulo e a albumina é secretada entre o casulo e a superfície clitelar. Então, o casulo e a albumina são levados a extremidade anterior por meio de ondas musculares e movimentos do corpo para trás. Durante esse deslocamento, o casulo recebe os óvulos e os espermatozóides (do parceiro), seguindo essa ordem. A fecundação ocorre na matriz da albumina, dentro do casulo. Os casulos são selados por contração de suas extremidades e são liberados pela extremidade anterior do corpo no substrato.
Os ovos dos oligoquetas são telolécitos, mas a quantidade de vitelo variará dependendo da quantidade de albumina secretada pois a quantidade de clitelo é inversamente proporcional à de albumina secretada.
O tempo de desenvolvimento varia de acordo com a espécie e as condições ambientais. O número de zigotos em um casulo vai de um a cerca de vinte.

3)Hirudinoidea:
3.1) Estrutura:
A maioria das cerca de 500 espécies dessa divisão são sanguesssugas verdadeiras. Como os oligoquetas, esses animais são clitelados. Possuem um número fixo de segmentos , que são tradicionalmente numerados com algarismos romanos, esses segmentos geralmente são cobertos por anelações superficiais, aparentando possuir muito mais segmentos.
No exterior, as sanguessuga são caracterizadas pelas ventosas anterior e posterior e pelo clitelo, não possuem parapódios ou certas em sua grande maioria. Internamente, o celoma é reduzido a uma série de canais intercomunicantes e espaços sem septos serialmente dispostos.
Os hiruidinóidos são achatados dorsiventralmente e divisível em cinco regiões. A região mais anterior é a cabeça e é composta pelo prostômio muito reduzido e pelos segmentos anteriores do corpo, um peristômio não é aparente, podendo estar ausente, e o prostômio pode estar fundido a alguns segmentos anteriores do corpo. A região anterior geralmente possui vários olhos e uma boca ventral circundada pela ventosa oral ou anterior. Os segmentos V-VIII formam a região pré-clitelar, e os segmentos IX-XI formam a região clitelar, a região pós clitelar é compreendida pelos segmentos XII-XXVII e a região posterior do corpo inclui a ventosa posterior dirigida ventralmente e é formada pelos segmentos XXVIII-XXXIV.

Glossiphoniidae leeches:encontrada em lagos e poças
3.2) Parede do corpo e celoma
A parede do corpo de uma sanguessuga é muito diferente se comparada a de outros anelídeos, principalmente devido a presença de uma camada de tecido conjuntivo dérmico abaixo da epiderme e peça redução do celoma.
Possuem uma cutícula fina sobre a camada única de células epidérmicas, a epiderme contém células glandulares muscosas. As musculaturas circular e longitudinal estão presentes mas estão organizadas mais esparsamente do que em poliquetas e oligoquetas, também estão presentes músculos dorsiventrais e músculos diagonais entre as camadas circular e longitudinal.
Há redução do celoma devido ao tecido conjuntivo abaixo da superfície corporal. Os espaços celômicos são representados por dirversos canais e espaços que aumentam o sistema circulatório nas sanguessugas rincobdélicas e o substituem completamente nas arrincobdélicas.
3.3) Sustentação e locomoção
O suporte se dá por uma construção corporal sólida, pelo tecido conjuntivo fibroso, bandas musculares e pelas propriedades hidrostáticas dos canais celômicos. O fato de não haverem compartimentos celômicos isolados, espaçosos e serialmente impossibilita alguns tipos de movimento conseguidos em poliquetas e oligoquetas.
Os músculos circulares e longitudinais agem antagonicamente contra um espaço interno funcionalmente único, cujo volume permanece constante.
Não possuindo cerdas ou parapódios, elas utilizam as ventosas como ponto de contato com o substrato. Os músculos circulares são contraídos fazendo com o que o único resultado possível desta ação é um alongamento de todo corpo e uma redução de seu diâmetro. Logo, o corpo é estendido para a frente e a ventosa anterior é aderida. Após esse movimento, a ventosa posterior se solta e os músculos longitudinais se contraem, encurtando o corpo e aumento o seu diâmetro, puxando a extremidade posterior para a frente. Esse mecanismo é conhecido como “mede-palmos”.
3.4) Alimentação
A maioria dos hirudíneos são ectoparasitas, e se alimentam de sangue ou fluidos corporais de seu hospedeiro. Grande parte é predadora de pequenos animais invertebrados, enquanto outra parte, é detritívora, se alimentando de animais mortos.
A obtenção dos alimentos envolve a estrutura da região anterior do trato digestivo, que inclui ou uma probóscide faríngea protraível, ou estruturas cortantes na forma de mandíbulas ou estiletes.
As duas subordens de Hirudinida são Rhynchobdellae e Arhyncobdellae. As sanguessugas Rhyncobdellae são caracterizadas pela presença de probóscide faríngea e ausência de mandíbula, já as Arhycobdellae não possuem probóscide faríngea e algumas poucas possuem mandíbula. No caso de probóscide ou mandíbula, a maioria das presas é engolida de uma vez e a maioria destes animais não é hospedeiro-específica, podendo se alimentar de outras maneiras se não aderida ao seu substrato.
Na hora da alimentação, a sanguessuga se prende ao hospedeiro pelas ventosas e pressiona a boca contra a superfície corporal do hospedeiro. Os animais que têm mandíbula causam uma incisão em forma de Y no hospedeiro. Músculos movem as mandíbulas para a frente e para trás, cortando a pele do hospedeiro, o animal libera anestésico enquanto faz suas incisões, libera hirudina dentro da ferida e então o sangue é sugado pela faringe muscular.
3.5) Sistema Digestivo
Os hirudíneos possuem sistema digestivo completo, formado pela região anterior estomodeal, região que inclui boca, mandíbulas, cavidade bucal, probóscide, faringe e esôfago, região mediana derivada da endoderme, região geralmente chamada de estômago ou papo e que porta grandes cercos que possuem capacidade de armazenamento, e pela região posterior proctodeal, em algumas sanguessugas essa região se difere da anterior pois um curto reto proctodeal conecta a região mediana do trato digestivo ao ânus, localizado dorsalmente, perto da junção do corpo à ventosa superior.
No caso das sanguessugas hematófagas, o estomodeu também contém massas de glândulas
salivares unicelulares que secretam hirudina nas sanguessugas hematófagas com mandíbula e produzem enzimas que auxiliam na penetração da probóscide, naquelas formas parasitas que não possuem mandíbulas. Não se sabe muito sobre digestão de hirudinóidos e o que se sabe são das sanguessugas hematófagas. As enzimas da região mediana do trato digestivo se limitam a exopeptidases, o que gera uma digestão extremamente lenta nesses animais e permite que esses animais fiquem longos períodos de tempo sem se alimentar.
A maioria das sanguessugas, predadoras ou parasitas, apresentam uma relação ecológica de mutualismo com bactérias, pois hospedam uma rica flora bacteriana intestinal, que auxiliam nos eventos digestivos e fornecem produtos metabólicos, como vitaminas, para o seu hospedeiro.
3.6) Circulação e trocas gasosas
A expansão da região do tecido conjuntivo está relacionada à redução do celoma e dos septos. Sem septos e mesentérios para compartimentalizar o celoma, a cavidade celômica tornou-se continua e ininterrupta ao longo do corpo das sanguessugas. Assim, o celoma desse grupo tornou-se o sistema circulatório, substituindo o papel o sistema hemal ancestral. Na maioria dos rincobdélicos, a circulação é uma combinação do sistema circulatório e dos espaços celômicos são reduzidos; nos arrincobdélicos, o sistema circulatório original foi substituído por outro inteiramente derivado do celoma reduzido. O fluido circulatório é movido através do sistema pela ação de vasos contráteis e pelos movimentos gerais do corpo.
As trocas gasosas são realizadas por difusão por meio das paredes do corpo. As brânquias ocorrem apenas nos ozobranquídeos e são projeções laterais da parede do corpo, foliáceas ou ramificadas. O pigmento respiratório, uma hemoglobina extracelular encontrada apenas nas sanguessugas arrincobdélicas, é responsável por cerca de metade do transporte de oxigênio.
3.7) Excreção e osmorregulação
As estruturas excretoras dos hirudinóideos são os nefrídios. Estas estruturas estão dispostas aos pares em cada metâmero, mas estão geralmente ausentes em vários segmentos anteriores e posteriores. Como resultado da redução do celoma e da perda dos septos intersegmentares, os túbulos nefridiais se aprofundaram no tecido conjuntivo e os nefróstomas se projetaram nos canais celômicos. Os nefróstomas são funis ciliados associados aos sistema circulatório celômico. Cada nefrídio leva a um nefridióporo ventrolateral.
O nefróstoma conduz a uma cápsula nefridial que é conectada a um duto composto por uma fileira de células através das quais corre um canal intracelular. Este canal conecta-se com uma curta câmara derivada de uma invaginação ectodérmica do nefridióporo. Em algumas sanguessugas verdadeiras, uma bexiga relativamente grande pode se formar perto do poro. A amônia é o principal excreta nitrogenado eliminado.
3.8) Curiosidades
Venenos ofídicos e de lepidópteros bem como salivas de sanguessugas, possuem um grande repertório de proteínas que interferem nos mecanismos de coagulação e fibrinólise. Têm se estudado enzimas fibrinolíticas, procoagulantes e anticoagulantes a partir destas fontes, além do mecanismo de ação destas proteínas com o intuito de estabelecer suas importâncias em processos de envenenamento e também com a finalidade de à partir delas desenharmos novos agentes terapêuticos. É um projeto foi proposto caracterizar os mecanismos de ação de diferentes fibrinolíticos de venenos ofídicos, do LOPAP (ativador de protrombina do veneno de Lonomia oblíqua) e do LEFAXIN (inibidor de FXa da sanguessuga Haementeria depressa), através do estudo de interações moleculares com suas proteínas alvo (fibrinogênio, fibrina, trombina, protrobina e FXa respectivamente) por técnicas de cinética em fluxo utilizando biocaptores no aparelho BIACORE. (AU) ( https://www.bv.fapesp.br/pt/auxilios/16390/proteinas-de-venenos-animais-que-interferem-nos-mecanismos-de-coagulacao-e-fibrinolise/
Saskatchewan
- Bibliografia:
Ruppert, E.E. & Barnes, R.D. 2005. Zoologia dos Invertebrados. 7ª Edição.
Brusca, R.C. &Brusca, G.J. 2007. Invertebrates. 2ª Edição.
De Oliveira, N.B., & Júnior, O.R.P. Venenos e peçonhas animais: aplicações tecnológicas e Biopirataria.
Alvaro E. Migotto. Poliqueto de escama. Banco de imagens Cifonauta. Disponível em: https://cifonauta.cebimar.usp.br/photo/10956/ Acesso em: 2013-11-12.
Alvaro E. Migotto. Larva de poliqueta. Banco de imagens Cifonauta. Disponível em: https://cifonauta.cebimar.usp.br/photo/10966/ Acesso em: 2013-11-12.
Alvaro E. Migotto. Poliqueta tubícola. Banco de imagens Cifonauta. Disponível em: https://cifonauta.cebimar.usp.br/photo/678/ Acesso em: 2013-11-12.
Alvaro E. Migotto. Poliqueta tubícola. Banco de imagens Cifonauta. Disponível em: https://cifonauta.cebimar.usp.br/photo/679/ Acesso em: 2013-11-12
Alvaro E. Migotto. Poliqueta terebelídeo. Banco de imagens Cifonauta. Disponível em: https://cifonauta.cebimar.usp.br/photo/11543/ Acesso em: 2013-11-12.
Alvaro E. Migotto. Verme-de-fogo. Banco de imagens Cifonauta. Disponível em: https://cifonauta.cebimar.usp.br/photo/6284/ Acesso em: 2013-11-12.
Alvaro E. Migotto. Poliqueta epítoco com massa de ovos presa ao ventre. Banco de imagens Cifonauta. Disponível em: https://cifonauta.cebimar.usp.br/photo/10905/ Acesso em: 2013-11-12
Alvaro E. Migotto. Poliqueto de escama. Banco de imagens Cifonauta. Disponível em: https://cifonauta.cebimar.usp.br/photo/10959/ Acesso em: 2013-11-12.
Lumbrineris sphaerocephala https://www.annelida.net/nz/Polychaeta/Family/Lumbrineridae/LumbriSphaerocephalaLY003.jpg Acesso em : 2013-11-12
{kind=link}
Macroinvertebrate Key Hirudinea https://extension.usu.edu/waterquality/htm/no-shell/macroinvertebrate-key-hirudinea Acesso em: 2013-11-12
Annelida- worm https://www.proprofs.com/flashcards/upload/a4163955.jpg Acesso em: 2013-11-12
{kind=link}
Annelida- worm https://www.proprofs.com/flashcards/upload/a4163949.jpg Acesso em: 2013-11-12
{kind=link}
Saskatchewan Leeches (Hirudinea) https://www.aquatax.ca/Leeches/broodingleech01.jpg Acesso em: 2013-11-12
{kind=link}
Earthworm dissection https://www.biologyjunction.com/images/earthw2.jpg Acesso em 2013-11-12
Earthworm dissection https://www.biologyjunction.com/images/earthw1.jpg Acesso em 2013-11-12
Earthworm dissection https://www.biologyjunction.com/images/clip0016.GIF Acesso em 2013-11-12